Breakthrough Achievement from Yeasen and Molefuture biotechnology
With the rapid development of biotechnology, mRNA therapy, as an emerging treatment method, has attracted much attention due to its strong programmability and fast response speed. In fields such as mRNA vaccines and gene therapy, efficient synthesis of high-quality mRNA is a crucial step in achieving therapeutic goals. In this process, T7 RNA polymerase plays a crucial role, as it can efficiently catalyze the in vitro synthesis of mRNA.
The dsRNA byproduct issue of T7 RNA polymerase
Although T7 RNA polymerase plays an important role in mRNA synthesis, the actual operation often results in the production of double-stranded RNA (dsRNA) as a byproduct. The presence of dsRNA not only reduces the yield and purity of mRNA but may also trigger nonspecific immune responses, affecting efficacy and safety. Therefore, reducing the production of dsRNA has become an urgent need in the industry. Currently, methods to reduce dsRNA mainly include optimizing reaction conditions and using auxiliary enzymes. Although these methods can reduce the production of dsRNA to some extent, they often come with issues such as complex operation and increased costs.
Why should we do T7 RNA polymerase engineering?
Directly modifying T7 RNA polymerase to reduce dsRNA production is a more fundamental solution. Enzyme-directed evolution techniques can improve its catalytic properties and increase the purity and yield of products by precisely changing the enzyme's amino acid sequence or structure. For T7 RNA polymerase, targeted modification can not only reduce dsRNA production but also enhance the overall efficiency and quality of mRNA synthesis.
As a supplier of mRNA in vitro synthesis raw materials, Yeasen Biotechnology, and its subsidiary Molefuture Biotechnology, have evolved a novel T7 RNA polymerase—utilizing the ZymeEditor directed evolution platform. In order to minimize the production of byproducts such as dsRNA during in vitro transcription processes, the evolution team has engineered T7 RNA polymerase through a combination of rational design and directed screening.
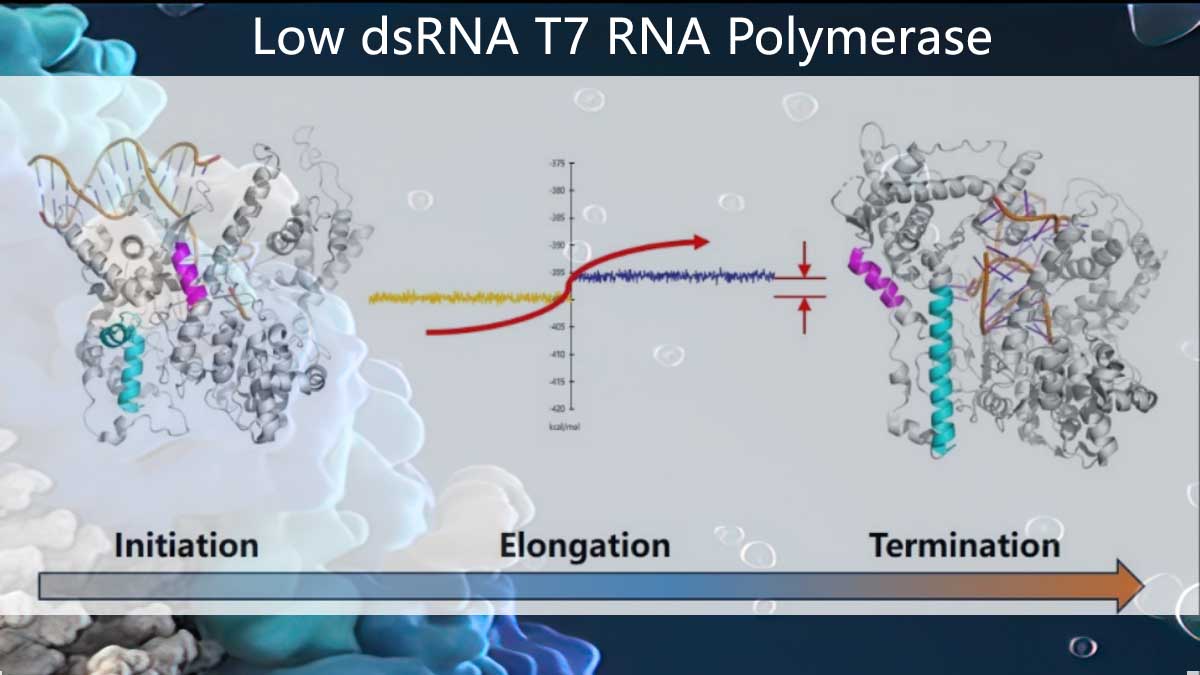
How does dsRNA generate?
According to literature reports, the production of the byproduct dsRNA mainly originates from two sources (Figure 1): The first is during the transition of T7 RNA polymerase from the initiation conformation to the elongation conformation in the early stages of transcription, the transcription ternary complex is prone to dissociation [1], resulting in the release of a large number of short RNA chains (Abortive RNA) with lengths around 20 nt into the system. These RNA chains act as "primers," complementary pairing with long RNA chains, and extend to produce dsRNA under the action of T7 RNA polymerase's RNA-dependent RNA polymerase activity (RDRP activity) [2,3]. The second source is triggered by the terminal ribonucleotide transfer activity possessed by T7 RNA polymerase. When the transcription reaction reaches the linear template's end, T7 RNA polymerase may randomly add several ribonucleotides without template dependence. These ribonucleotides, forming "random primers," may reverse-fold and complementarily pair with the target mRNA region, extending to produce dsRNA. The dsRNA produced through looping is called loopback dsRNA [3,4]. Therefore, in order to reduce dsRNA byproducts, the focus of T7 RNA polymerase modification lies in stabilizing the early transcription ternary complex or weakening the terminal transfer activity and RNA dependent RNA Polymerase (RDRP) activity of T7 RNA polymerase.

Figure 1. Schematic diagram of energy barrier and dsRNA formation principle (unpublished data)
How to overcome energy barrier of T7 RNA polymerase conformational transition?
Through structural and free energy analysis, the team, on one hand, discovered that along with the conformational changes of T7 RNA polymerase, there is also a change in energy. The free energy of the elongation conformation is significantly higher than that of the initiation conformation, indicating that the transcription process needs to overcome a higher energy barrier to produce complete mRNA chains. A high energy barrier is unfavorable for smooth conformational transition. Therefore, the team utilized rational screening methods to identify over 20 hotspot amino acids and constructed corresponding saturation mutagenesis libraries.
How to modify the terminal transfer and RDRP activities of T7 RNA polymerase
On the other hand, considering the limited reports on the functions, sites, and structural domains related to the terminal transfer and RDRP activities of T7 RNA polymerase, and because these two activities, which are functionally similar, likely share key sites with the main reaction of T7 RNA polymerase—transcription activity (i.e., DNA-dependent RNA polymerization activity, DDRP)—it is difficult to find hotspot amino acids for site-directed modification. Therefore, the team simultaneously constructed several random mutation libraries based on error-prone PCR, combined with the high-throughput evolution capacity of the ZymeEditor platform, resulting in a diversity exceeding 10^6 for each individual library.
Probe based discovery of ideal T7 RNA polymerase
To balance the production of T7 RNA polymerase and monitor the content of dsRNA in the in vitro transcription reaction, the team, with reference to optimized transcription templates, carefully designed multiple Fluorescent probes targeting different regions of the target mRNA products. These probes associate information such as reaction yield, integrity, and dsRNA content within the system with fluorescence signals. The stronger the fluorescence intensity of the system, the more advantageous the performance of the mutant. In theory, this screening method can rank mutants based on the fluorescence intensity of different probes, obtaining T7 RNA polymerase mutants with high yield or reduced Abortive RNA, as well as mutants with improved integrity or reduced loopback dsRNA. When the reaction template is replaced with RNA, mutants with reduced RDRP activity can also be negatively screened, improving the template selectivity of T7 RNA polymerase. Additionally, by adding modified nucleotides, cap analogs, and increasing the reaction temperature in the droplet reaction system, further screening can be conducted to select T7 RNA polymerase mutants that tolerate non-natural substrates and exhibit improved thermal stability.
How to screen best T7 RNA polymerase?
Based on the size of the libraries, the team developed ultra-high-throughput screening processes based on fluorescence-activated droplet sorting (FADS) and high-throughput screening processes based on traditional microplate methods (MTPS). Through multiple rounds of experiments, over 10^7 mutants were screened in total, resulting in multiple mutants with significantly reduced dsRNA content. Among them, some mutants showed a decrease in dsRNA caused by Abortive RNA in the reaction, suggesting that the elongation conformation of these polymerase mutants is more stable. Some mutants showed a decrease in loopback dsRNA content in the reaction, indicating a weakening of terminal transfer activity or RDRP activity in these mutants.
Given that the performance advantages of the aforementioned mutants stem from different mechanisms, the team has constructed DNA shuffling libraries targeting them. After screening, the team finally obtained superior-performing mutants with extreme low dsRNA generation while not affecting the yields and mRNA integrity (Figure 2). However,the yields of a mutant G47A+884G discovered by Moderna was affected[5] (unpublished).

Figure 2. Enzyme evolution process and characterization of mutant performance
(unpublished)
Analysis of dsRNA generation by T7 RNA polymerase variants?
After quantitative analysis, as shown in Figure 3, using a 9 kb transcription template under co-transcription capping conditions, the wild-type T7 RNA polymerase produced approximately 2.9 ng/μg of dsRNA when using natural nucleotides, while a commercial T7 enzyme produced about 0.18 ng/μg of dsRNA. However, the dsRNA production of each T7 RNA polymerase variant constructed was less than 0.10 ng/μg. Particularly, one variant was only 0.015 ng/μg, nearly 200 times lower than the wild type and 12 times lower than the commercial product. After repeated testing, the performance advantage of the variants in reducing dsRNA was consistently observed under different reaction conditions, indicating good system compatibility of these mutants and laying a foundation for further reducing dsRNA production through reaction buffer optimization. In addition, FADS screening also obtained multiple variants with improved product integrity and thermal stability, which can meet the requirements of a wider range of in vitro transcription applications

Figure 3. Evaluation of dsRNA generation by T7 RNA polymerase variants assessed using the Yeasen Double-stranded RNA (dsRNA) ELISA Kit and J2 antibody dot blot analysis.
At present, several partners have already tried the T7 RNA polymerase variants and we have received high praise from them. The use of the new enzyme simplifies the mRNA purification process, enhances work efficiency, and demonstrates outstanding performance in multiple application scenarios.
About Molefuture Biotechnology
Molefuture Biotechnology is a subsidiary of Yeasen Biotechnology (Shanghai) Co., Ltd., specializing in providing customized enzyme modification solutions driven by enzyme evolution technology. Leveraging the resources of Yeasen Biotechnology, Molefuture operates on six major technological platforms: the ZymeEditor innovative enzyme evolution platform, a multi-host high-efficiency expression platform, fermentation process development platform, purification process development platform, ultra-clean enzyme production platform, and enzyme analysis and quality control platform. With these six technological platforms, Molefuture can offer a complete set of customized solutions including enzyme-directed evolution, new enzyme development, enzyme process development, and GMP-level scale-up production to meet the application demands of enzymes in areas such as in vitro diagnostics, biomedicine, synthetic biology, medical aesthetics, pharmaceutical intermediates, et. al.
Ordering Information
The following are representative products offered by Yeasen. Additional sizes are available. Our products are highly optimized to work in concert, to help ensure superior performance and reproducibility. We can also provide customized services. If you’re interested in a product that isn’t shown, contact us and we’ll work with you to meet your needs.
Regarding reading:
References: